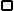Аминная гипотеза депрессии. В начале 50-х гг XX века после внедрения резерпина в широкую клиническую практику стало ясно, что, вызывая опустошение нейрональных депо аминных нейромедиаторов, резерпин индуцирует ятрогенную депрессию. Был сделан кардинальный вывод – депрессия связана со снижением функции аминных медиаторных систем мозга. Именно этот вывод и был положен в основу т.н. аминной гипотезы депрессии.

На нейронах эмоциогенных зон ретикулярной формации сходятся адренергические и серотонинергические нервные окончания. Высвобождающийся из адренергических окончаний норадреналин влияет на постсинаптические a1 и b-адренорецепторы:
· a1-адренорецепторы – низкоаффинные, реагируют только на значительные (физиологические) концентрации норадреналина и в норме тормозят электрическую активность нейрона.
· b-адренорецепторы – высокоаффинные, реагируют даже на низкие (субфизиологические) концентрации норадреналина и усиливают электрическую активность нейрона.
Высвобождающийся из серотонинергических окончаний серотонин влияет на постсинаптические 5-НТ1А, 5-НТ2 и 5-НТ3 типы рецепторов:
· 5-НТ1А-рецепторы – высокоаффинные, реагируют на низкие (субфизиологические) концентрации серотонина и тормозят электрическую активность нейрона.
· 5-НТ2 и 5-НТ3 типы рецепторов – низкоаффинные, реагируют только на значительные (физиологические) концентрации серотонина и усиливают электрическую активность нейрона.
Кроме постсинаптических влияний моноаминергические терминали нейронов посылают взаимные тормозящие коллатерали друг к другу, которые взаимно ограничивают секрецию медиаторов через a2 и 5-НТ1D-типы рецепторов.
В обычных условиях нейроны выделяют достаточно медиатора для активации низкоаффинных рецепторов – сочетание стимулирующих влияний 5-НТ2/5-НТ3 рецепторов серотонина и тормозных влияний a1-адренорецепторов нейтрализует друг друга и на выходе воспринимающего нейрона формируется постоянный ритмичный сигнал. Этому соответствует ровный обычный фон настроения.
Дефицит серотонина приводит к тому, что низкоаффинные 5-НТ2/5-НТ3 рецепторы уже не могут активироваться и на первый план выступают тормозящие нейрон высокоаффинные 5-НТ1А-рецепторы. Их эффект накладывается на эффект тормозящих нейрон a1-адренорецепторов и, в конечном итоге, нейрон генерирует редкие, нерегулярные сигналы сопровождающиеся возникновением апатии, заторможенности и астенически-адинамической формы депрессии. Кроме того, возникает порочный круг, т.к. в условиях недостатка серотонина прекращается торможение адренергических нейронов по коллатералям через 5-НТ1D-тип рецепторов и выброс норадреналина возрастает, с вовлечением в патологический процесс все новых и новых низкоаффинных a1-адренорецепторов.
Дефицит норадреналина сопровождается потерей чувствительности a1-адренорецепторов и преобладанием эффекта высокоаффинных b-адренорецепторов, которые повышают активность воспринимающего нейрона. Эффект b-адренорецепторов суммируется с эффектом серотониновых 5-НТ2/5-НТ3 рецепторов и импульсация эффекторного нейрона резко возрастает. Одновременно, в условиях дефицита норадреналина, прекращается торможение активности серотонинергических нейронов коллатеральными веточками с a2-адренорецепторами. Такое чрезмерное усиление ответа нейрона приводит к развитию тревожно-ажитированной формы депрессии.




.gif)